|
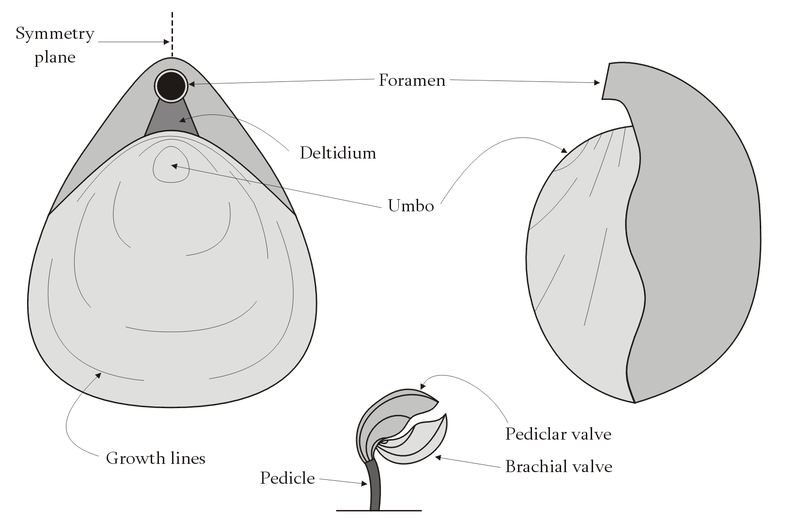
Brachiopods have a mantle
that secretes and lines
the shell, and encloses the internal organs. The body occupies typically
about one-third of the internal space inside the shell, nearest the hinge.
The rest of the mantle encloses a water-filled space containing the
lophophore, a crown of tentacles that
filters food
particles out of the water. In all species the lophophore is supported by
cartilage and by a
hydrostatic skeleton.
The
lophophore
filters food, mostly
phytoplankton, out of
the water. From there the food is transported in succession to: the grooves
along the bases of the lophophore's tentacles; the mouth;
pharynx;
oesophagus; and
finally the stomach, where the
food is digested. Nutrients
are transported from the stomach throughout the
coelom
(main body cavity), include the mantle lobes, by cilia. The wastes produced
by
metabolism are broken
into
ammonia, which is
eliminated by
diffusion through the
mantle and lophophore.
The lophophore and mantle are the only surfaces that absorb
oxygen
and eliminate
carbon dioxide.
Oxygen seems to be distributed by the fluid of the coelom. The heart is
above the stomach, and the blood vessels connect it to the major organs.
However, the main function of the blood may be to deliver nutrients. The
maximum oxygen consumption of brachiopods is low, and their minimum
requirement is not measurable.
The "brain" of adult
articulates consists of two
ganglia, one above and the other below
 |
|
Lingula anatina from Stradbroke Island, Australia.
|
the oesophagus. Adult inarticulates have only the lower ganglion. Nerves
run to the lophophore, the mantle lobes and the muscles that operate the
valves. Many brachiopods close their valves if shadows appear above them,
but the cells responsible for this are unknown. Some brachiopods have
statocysts which detect changes in the
animals'
balance.
Lifespans range from 3 to over 30 years. Ripe
gametes
(ova
or
sperm) float from the
gonads into the main coelom and then exit
into the mantle cavity. The
larvae
of inarticulate brachiopods are miniature adults, with lophophores that
enable the larvae to feed and swimming for months, until the animals become
heavy enough to settle to the seabed. Larvae of articulate species are
different from the adult forms, live only on
yolk,
remain only among the plankton for only a few days, and then
metamorphosing.
In addition to the traditional classification into inarticulate and
articulate brachiopods, two approaches appeared in the 1990s: grouping the
inarticulate
Craniida with
articulate brachiopods, as both used the same material in the
mineral
layers of the shell; and making the Craniida a third group, as their outer
organic layer is
different from that of either the others. However, some
taxonomists believe it is premature to
suggest higher levels of classification such as
order, and recommend
a bottom-up approach that identifies genera and then groups these into
intermediate groups. Traditionally brachiopods have been regarded as members
of or as a
sister group to the
deuterostomes, a
super-phylum which includes
chordates and
echinoderms. One type of analysis of
brachiopods' evolutionary relationships has always placed brachiopods as
protostomes, while
another type has split between placing brachiopods among the protostomes or
the deuterostomes.
In 2003 it was suggested that brachiopods evolved from an ancestor similar
to
Halkieria, a
slug-like
animal with "chain
mail" on its back and a shell at the front
and rear end, and that the ancestral brachiopod converted its shells into a
pair of valves by folding the rear part of its body under its front.
However, new fossils found in 2007 to 2008 showed that
tommotiids' "chain mail" formed the tube of
a
sessile animal, and
that one resembled
phoronids, which are
close relatives or a sub-group of brachiopod, while the other tommotiid
bearing two symmetrical plates that might be an early form of brachiopod
valves. Lineages that have both fossil and
extant brachiopods appeared in the early
Cambrian,
Ordovician and
Carboniferous
periods respectively.
Other lineages have arisen and then become extinct, sometimes during severe
mass extinctions. At
their peak in the
Paleozoic
era the brachiopods were among the most
abundant filter-feeders and reef-builders, and occupied other
ecological niches,
including swimming in the jet-propulsion style of
scallops. Brachiopod fossils have been
useful indicators of climate changes during the
Paleozoic era. However, after the Permian–Triassic extinction event,
brachiopods recovered only a third of their former diversity. A study in
2007 concluded that brachiopods were especially vulnerable to the
Permian–Triassic extinction, as they built calcareous hard parts (made of
calcium carbonate)
and had low
metabolic rates and
weak respiratory systems. It was often thought that brachiopods were in
decline after the Permian–Triassic extinction, and were out-competed by
bivalves. However, a study in 1980 concluded that: both brachiopods and
bivalves increased all the way from the Paleozoic to modern times, but
bivalves increased faster; after the Permian–Triassic extinction,
brachiopods for the first time were less diverse than bivalves.
Brachiopods live only in the
sea, and most species avoid locations with strong currents or waves.
Articulate species have larvae that settling quickly and form dense
populations in well-defined areas,
while inarticulate larvae swimming for up to a month and have wide ranges.
Brachiopods now live mainly in cold and low-light conditions. Fish and
crustaceans seem to find brachiopod flesh distasteful and seldom attack
them. Among brachiopods only the lingulids have been fished commercially, on
a very small scale. One brachiopod species may be a measure of environmental
conditions around an oil terminal being built in Russia on the shore of the
Sea of Japan.
Fossil Record
 |
|
Above:
A dense assemblage of the Ordovician species
Dalmanella meeki
|
Below:
Rhynchotrema dentatum, a rhynchonellid brachiopod
from the Cincinnatian (Upper Ordovician) of southeastern Indiana.
|
 |
Over
12,000 fossil species are recognized,grouped
into over 5,000 genera.
While the largest modern brachiopods are 100 millimetres (3.9 in) long,
a few fossils measure up to 200 millimetres (7.9 in) wide.
The earliest confirmed brachiopods have been found in the early Cambrian,
inarticulate forms appearing first, followed soon after by articulate forms.
Three unmineralized species have also been found in the Cambrian, and
apparently represent two distinct groups that evolved from mineralized
ancestors. The inarticulate
Lingula is often
called a "living
fossil", as very similar
genera
have been all the way back to the Ordovician. On the other hand, articulate
brachiopods have produced major diversifications, and severe mass
extinctions – but the articulate Rhynchonellida and Terebratulida, the most
diverse present-day groups, appeared at the start of the Ordovician and
Carboniferous respectively.
At their
peak in the
Paleozoic the
brachiopods were among the most abundant filter-feeders and reef-builders,
and occupied other
ecological niches,
including swimming in the jet-propulsion style of
scallops. However, after the
Permian–Triassic extinction event, informally known as the "Great Dying",
brachiopods recovered only a third of their former diversity.
It was often thought that brachiopods were actually declining in diversity,
and that in some way bivalves out-competed them. However, in 1980 studies
produced a statistical analysis that concluded that: both brachiopods and
bivalves increased all the way from the Paleozoic to modern times, but
bivalves increased faster; the Permian–Triassic extinction was moderately
severe for bivalves but devastating for brachiopods, so that brachiopods for
the first time were less diverse than bivalves and their diversity after the
Permian increased from a very low base; there is no evidence that bivalves
out-competed brachiopods, and short-term increases or decreases for both
groups appeared at the same times.
Brachiopod fossils have been
useful indicators of climate changes during the
Paleozoic era. When global temperatures
were low, as in much of the Ordovician, the large difference in temperatures
between equator and poles created different collections of fossils at
different latitudes. On the other hand warmer periods, such much of the
Silurian, created smaller difference in temperatures, and all seas at the
low to middle latitudes were colonized by the same few brachiopod species.
End of Reading

Return to the
Old Earth Ministries Online Earth
History Curriculum homepage.
Source:
Brachiopod
|