 |
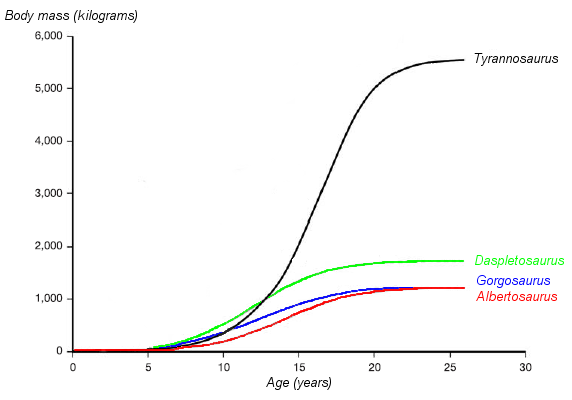
| A graph showing the hypothesized growth curve,
body mass versus age (drawn in black, with other tyrannosaurids for
comparison). VIDEO T Rex Maturation |
Histology has also allowed the age of other specimens to be determined. Growth curves can be developed when the ages of different specimens are plotted on a graph along with their mass. A Tyrannosaurus rex growth curve is S-shaped, with juveniles remaining under 1,800 kg (4,000 lb) until approximately 14 years of age, when body size began to increase dramatically. During this rapid growth phase, a young Tyrannosaurus rex would gain an average of 600 kg (1,300 lb) a year for the next four years. At 18 years of age, the curve plateaus again, indicating that growth slowed dramatically. For example, only 600 kg (1,300 lb) separated the 28-year-old "Sue" from a 22-year-old Canadian specimen (RTMP 81.12.1). Another recent histological study performed by different workers corroborates these results, finding that rapid growth began to slow at around 16 years of age. This sudden change in growth rate may indicate physical maturity, a hypothesis which is supported by the discovery of medullary tissue in the femur of a 16 to 20-year-old Tyrannosaurus rex from Montana (MOR 1125, also known as "B-rex"). Medullary tissue is found only in female birds during ovulation, indicating that "B-rex" was of reproductive age. Further study indicates an age of 18 for this specimen. Other tyrannosaurids exhibit extremely similar growth curves, although with lower growth rates corresponding to their lower adult sizes.
Over half of the known Tyrannosaurus rex specimens appear to have died within six years of reaching sexual maturity, a pattern which is also seen in other tyrannosaurs and in some large, long-lived birds and mammals today. These species are characterized by high infant mortality rates, followed by relatively low mortality among juveniles. Mortality increases again following sexual maturity, partly due to the stresses of reproduction. One study suggests that the rarity of juvenile Tyrannosaurus rex fossils is due in part to low juvenile mortality rates; the animals were not dying in large numbers at these ages, and so were not often fossilized. However, this rarity may also be due to the incompleteness of the fossil record or to the bias of fossil collectors towards larger, more spectacular specimens.
Sexual Dimorphism
As the number of specimens increased, scientists began to analyze the variation between individuals and discovered what appeared to be two distinct body types, or morphs, similar to some other theropod species. As one of these morphs was more solidly built, it was termed the 'robust' morph while the other was termed 'gracile.' Several morphological differences associated with the two morphs were used to analyze sexual dimorphism in Tyrannosaurus rex, with the 'robust' morph usually suggested to be female. For example, the pelvis of several 'robust' specimens seemed to be wider, perhaps to allow the passage of eggs. It was also thought that the 'robust' morphology correlated with a reduced chevron on the first tail vertebra, also ostensibly to allow eggs to pass out of the reproductive tract, as had been erroneously reported for crocodiles.
In recent years, evidence for sexual dimorphism has been weakened. A 2005 study reported that previous claims of sexual dimorphism in crocodile chevron anatomy were in error, casting doubt on the existence of similar dimorphism between Tyrannosaurus rex genders. A full-sized chevron was discovered on the first tail vertebra of "Sue," an extremely robust individual, indicating that this feature could not be used to differentiate the two morphs anyway. As Tyrannosaurus rex specimens have been found from Saskatchewan to New Mexico, differences between individuals may be indicative of geographic variation rather than sexual dimorphism. The differences could also be age-related, with 'robust' individuals being older animals.
Only a single Tyrannosaurus rex specimen has been conclusively shown to belong to a specific gender. Examination of "B-rex" demonstrated the preservation of soft tissue within several bones. Some of this tissue has been identified as a medullary tissue, a specialized tissue grown only in modern birds as a source of calcium for the production of eggshell during ovulation. As only female birds lay eggs, medullary tissue is only found naturally in females, although males are capable of producing it when injected with female reproductive hormones like estrogen. This strongly suggests that "B-rex" was female, and that she died during ovulation. Recent research has shown that medullary tissue is never found in crocodiles, which are thought to be the closest living relatives of dinosaurs, aside from birds. The shared presence of medullary tissue in birds and theropod dinosaurs is further evidence of the close evolutionary relationship between the two.
Posture
Like many bipedal dinosaurs, Tyrannosaurus rex was historically depicted as a 'living tripod', with the body at 45 degrees or less from the vertical and the tail dragging along the ground, similar to a kangaroo. This concept dates from Joseph Leidy's 1865 reconstruction of Hadrosaurus, the first to depict a dinosaur in a bipedal posture. Henry Fairfield Osborn, former president of the American Museum of Natural History (AMNH) in New York City, who believed the creature stood upright, further reinforced the notion after unveiling the first complete Tyrannosaurus rex skeleton in 1915. It stood in this upright pose for nearly a century, until it was dismantled in 1992. By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the dislocation or weakening of several joints, including the hips and the articulation between the head and the spinal column. The inaccurate AMNH mount inspired similar depictions in many films and paintings (such as Rudolph Zallinger's famous mural The Age Of Reptiles in Yale University's Peabody Museum of Natural History) until the 1990s, when films such as Jurassic Park introduced a more accurate posture to the general public. Modern representations in museums, art, and film show Tyrannosaurus rex with its body approximately parallel to the ground and tail extended behind the body to balance the head.
Arms
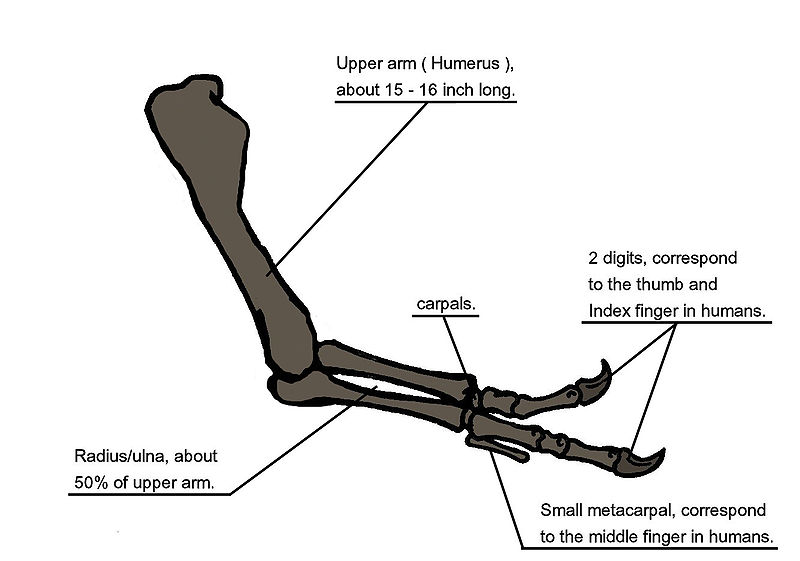 |
| Above: Arm Anatomy Below: Closeup of forelimb (Source) |
 |
When Tyrannosaurus rex was first discovered, the humerus was the only element of the forelimb known. For the initial mounted skeleton as seen by the public in 1915, Osborn substituted longer, three-fingered forelimbs like those of Allosaurus. However, a year earlier, Lawrence Lambe described the short, two-fingered forelimbs of the closely related Gorgosaurus. This strongly suggested that Tyrannosaurus rex had similar forelimbs, but this hypothesis was not confirmed until the first complete Tyrannosaurus rex forelimbs were identified in 1989, belonging to MOR 555 (the "Wankel rex"). The remains of "Sue" also include complete forelimbs. Tyrannosaurus rex arms are very small relative to overall body size, measuring only 1 metre (3.3 ft) long. However, they are not vestigial but instead show large areas for muscle attachment, indicating considerable strength. It has been suggested that the forelimbs were used to assist the animal in rising from a prone position. Another possibility is that the forelimbs held struggling prey while it was dispatched by the tyrannosaur's enormous jaws. This hypothesis may be supported by biomechanical analysis. Tyrannosaurus rex forelimb bones exhibit extremely thick cortical bone, indicating that they were developed to withstand heavy loads. The biceps brachii muscle of a full-grown Tyrannosaurus rex was capable of lifting 199 kilograms (439 lb) by itself; this number would only increase with other muscles (like the brachialis) acting in concert with the biceps. A Tyrannosaurus rex forearm also had a reduced range of motion, with the shoulder and elbow joints allowing only 40 and 45 degrees of motion, respectively. In contrast, the same two joints in Deinonychus allow up to 88 and 130 degrees of motion, respectively, while a human arm can rotate 360 degrees at the shoulder and move through 165 degrees at the elbow. The heavy build of the arm bones, extreme strength of the muscles, and limited range of motion may indicate a system designed to hold fast despite the stresses of a struggling prey animal.
Skin and Feathers
In 2004, the scientific journal Nature published a report describing an early tyrannosauroid, Dilong paradoxus, from the famous Yixian Formation of China. As with many other theropods discovered in the Yixian, the fossil skeleton was preserved with a coat of filamentous structures which are commonly recognized as the precursors of feathers. It has also been proposed that Tyrannosaurus and other closely related tyrannosaurids had such protofeathers. However, skin impressions from large tyrannosaurid specimens show mosaic scales. While it is possible that protofeathers existed on parts of the body which have not been preserved, a lack of insulatory body covering is consistent with modern multi-ton mammals such as elephants, hippopotamus, and most species of rhinoceros. As an object increases in size, its ability to retain heat increases due to its decreasing surface area-to-volume ratio. Therefore, as large animals evolve in or disperse into warm climates, a coat of fur or feathers loses its selective advantage for thermal insulation and can instead become a disadvantage, as the insulation traps excess heat inside the body, possibly overheating the animal. Protofeathers may also have been secondarily lost during the evolution of large tyrannosaurids like Tyrannosaurus, especially in warm Cretaceous climates.
Thermoregulation
Tyrannosaurus, like most dinosaurs, was long thought to have an ectothermic ("cold-blooded") reptilian metabolism. The idea of dinosaur ectothermy was challenged by scientists like Robert T. Bakker and John Ostrom in the early years of the "Dinosaur Renaissance", beginning in the late 1960s. Tyrannosaurus rex itself was claimed to have been endothermic ("warm-blooded"), implying a very active lifestyle. Since then, several paleontologists have sought to determine the ability of Tyrannosaurus to regulate its body temperature. Histological evidence of high growth rates in young Tyrannosaurus rex, comparable to those of mammals and birds, may support the hypothesis of a high metabolism. Growth curves indicate that, as in mammals and birds, Tyrannosaurus rex growth was limited mostly to immature animals, rather than the indeterminate growth seen in most other vertebrates.
Oxygen isotope ratios in fossilized bone are sometimes used to determine the temperature at which the bone was deposited, as the ratio between certain isotopes correlates with temperature. In one specimen, the isotope ratios in bones from different parts of the body indicated a temperature difference of no more than 4 to 5°C (7 to 9°F) between the vertebrae of the torso and the tibia of the lower leg. This small temperature range between the body core and the extremities was claimed by paleontologist Reese Barrick and geochemist William Showers to indicate that Tyrannosaurus rex maintained a constant internal body temperature (homeothermy) and that it enjoyed a metabolism somewhere between ectothermic reptiles and endothermic mammals. Other scientists have pointed out that the ratio of oxygen isotopes in the fossils today does not necessarily represent the same ratio in the distant past, and may have been altered during or after fossilization (diagenesis). Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus). Ornithischian dinosaurs also showed evidence of homeothermy, while varanid lizards from the same formation did not. Even if Tyrannosaurus rex does exhibit evidence of homeothermy, it does not necessarily mean that it was endothermic. Such thermoregulation may also be explained by gigantothermy, as in some living sea turtles.
Locomotion
There are two main issues concerning the locomotory abilities of Tyrannosaurus: how well it could turn; and what its maximum straight-line speed was likely to have been. Both are relevant to the debate about whether it was a hunter or a scavenger (see below).
Tyrannosaurus may have been slow to turn, possibly taking one to two seconds to turn only 45° — an amount that humans, being vertically oriented and tail-less, can spin in a fraction of a second. The cause of the difficulty is rotational inertia, since much of Tyrannosaurus’ mass was some distance from its center of gravity, like a human carrying a heavy timber — although it might have reduced the average distance by arching its back and tail and pulling its head and forelimbs close to its body, rather like the way ice skaters pull their arms closer in order to spin faster.
Scientists have produced a wide range of maximum speed estimates, mostly around 11 meters per second (40 km/h; 25 mph), but a few as low as 5–11 meters per second (18–40 km/h; 11–25 mph), and a few as high as 20 meters per second (72 km/h; 45 mph). Researchers have to rely on various estimating techniques because, while there are many tracks of very large theropods walking, so far none have been found of very large theropods running—and this absence may indicate that they did not run. Scientists who think that Tyrannosaurus was able to run point out that hollow bones and other features that would have lightened its body may have kept adult weight to a mere 4.5 metric tons (5.0 short tons) or so, or that other animals like ostriches and horses with long, flexible legs are able to achieve high speeds through slower but longer strides. Additionally, some have argued that Tyrannosaurus had relatively larger leg muscles than any animal alive today, which could have enabled fast running 40–70 kilometers per hour (25–43 mph).
Jack Horner and Don Lessem argued in 1993 that Tyrannosaurus was slow and probably could not run (no airborne phase in mid-stride), because its ratio of femur (thigh bone) to tibia (shin bone) length was greater than 1, as in most large theropods and like a modern elephant. However, Holtz (1998) noted that tyrannosaurids and some closely related groups had significantly longer distal hindlimb components (shin plus foot plus toes) relative to the femur length than most other theropods), and that tyrannosaurids and their close relatives had a tightly interlocked metatarsus that more effectively transmitted locomotory forces from the foot to the lower leg than in earlier theropods ("metatarsus" means the foot bones, which function as part of the leg in digitigrade animals). He therefore concluded that tyrannosaurids and their close relatives were the fastest large theropods.
Christiansen (1998) estimated that the leg bones of Tyrannosaurus were not significantly stronger than those of elephants, which are relatively limited in their top speed and never actually run (there is no airborne phase), and hence proposed that the dinosaur's maximum speed would have been about 11 meters per second (40 km/h; 25 mph), which is about the speed of a human sprinter. But he also noted that such estimates depend on many dubious assumptions.
Farlow and colleagues (1995) have argued that a Tyrannosaurus weighing 5.4 metric tons (6.0 short tons) to 7.3 metric tons (8.0 short tons) would have been critically or even fatally injured if it had fallen while moving quickly, since its torso would have slammed into the ground at a deceleration of 6 g (six times the acceleration due to gravity, or about 60 meters/s²) and its tiny arms could not have reduced the impact. However, giraffes have been known to gallop at 50 kilometers per hour (31 mph), despite the risk that they might break a leg or worse, which can be fatal even in a "safe" environment such as a zoo. Thus it is quite possible that Tyrannosaurus also moved fast when necessary and had to accept such risks.
Most recent research on Tyrannosaurus locomotion does not support speeds faster than 40 kilometers per hour (25 mph), i.e. moderate-speed running. For example, a 2002 paper in the journal Nature used a mathematical model (validated by applying it to three living animals, alligators, chickens, and humans; additionally later eight more species including emus and ostriches) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph). They found that proposed top speeds in excess of 40 kilometers per hour (25 mph) were unfeasible, because they would require very large leg muscles (more than approximately 40–86% of total body mass). Even moderately fast speeds would have required large leg muscles. This discussion is difficult to resolve, as it is unknown how large the leg muscles actually were in Tyrannosaurus. If they were smaller, only 18 kilometers per hour (11 mph) walking/jogging might have been possible.
A study in 2007 used computer models to estimate running speeds, based on data taken directly from fossils, and claimed that Tyrannosaurus rex had a top running speed of 8 meters per second (29 km/h; 18 mph). An average professional soccer player would be slightly slower, while a human sprinter can reach 12 meters per second (43 km/h; 27 mph). Note that these computer models predict a top speed of 17.8 meters per second (64 km/h; 40 mph) for a 3-kilogram (6.6 lb) Compsognathus (probably a juvenile individual).
Those who argue that Tyrannosaurus was incapable of running estimate the top speed of Tyrannosaurus at about 17 kilometers per hour (11 mph). This is still faster than its most likely prey species, hadrosaurids and ceratopsians. In addition, some advocates of the idea that Tyrannosaurus was a predator claim that tyrannosaur running speed is not important, since it may have been slow but still faster than its probable prey. However, Paul and Christiansen (2000) argued that at least the later ceratopsians had upright forelimbs and the larger species may have been as fast as rhinos. Healed Tyrannosaurus bite wounds on ceratopsian fossils are interpreted as evidence of attacks on living ceratopsians (see below). If the ceratopsians that lived alongside Tyrannosaurus were fast, that casts doubt on the argument that Tyrannosaurus did not have to be fast to catch its prey.
Feeding Strategies
The debate about whether Tyrannosaurus was a predator or a pure scavenger is as old as the debate about its locomotion. Ever since the first discovery of Tyrannosaurus most scientists have agreed that it was a predator, although like modern large predators it would have been happy to scavenge or steal another predator's kill if it had the opportunity.
Noted hadrosaur expert Jack Horner is currently the major advocate of the idea that Tyrannosaurus was exclusively a scavenger and did not engage in active hunting at all. Horner has presented several arguments to support the pure scavenger hypothesis:
Tyrannosaur arms are short when compared to other known predators. Horner argues that the arms were too short to make the necessary gripping force to hold on to prey.
Tyrannosaurs had large olfactory bulbs and olfactory nerves (relative to their brain size). These suggest a highly developed sense of smell which could sniff out carcasses over great distances, as modern vultures do. Research on the olfactory bulbs of dinosaurs has shown that Tyrannosaurus had the most highly developed sense of smell of 21 sampled dinosaurs. Opponents of the pure scavenger hypothesis have used the example of vultures in the opposite way, arguing that the scavenger hypothesis is implausible because the only modern pure scavengers are large gliding birds, which use their keen senses and energy-efficient gliding to cover vast areas economically. However, researchers from Glasgow concluded that an ecosystem as productive as the current Serengeti would provide sufficient carrion for a large theropod scavenger, although the theropod might have had to be cold-blooded in order to get more calories from carrion than it spent on foraging (see Warm-bloodedness of dinosaurs). They also suggested that modern ecosystems like Serengeti have no large terrestrial scavengers because gliding birds now do the job much more efficiently, while large theropods did not face competition for the scavenger ecological niche from gliding birds.
Tyrannosaur teeth could crush bone, and therefore could extract as much food (bone marrow) as possible from carcass remnants, usually the least nutritious parts. Karen Chin and colleagues have found bone fragments in coprolites (fossilized dung) that they attribute to tyrannosaurs, but point out that a tyrannosaur's teeth were not well adapted to systematically chewing bone like hyenas do to extract marrow.
Since at least some of Tyrannosaurus's potential prey could move quickly, evidence that it walked instead of ran could indicate that it was a scavenger. On the other hand, recent analyses suggest that Tyrannosaurus, while slower than large modern terrestrial predators, may well have been fast enough to prey on large hadrosaurs and ceratopsians
